什么是组蛋白?
DNA就像生命的一个神奇、错综复杂的缠绕之绳,它的魔法在于与组蛋白的协同作用,将两米长的DNA魔法般地压缩在一个微小的6微米细胞内,储存着遗传信息的秘密。
1. 什么是组蛋白?
组蛋白于1884年首次被Albrecht Kossel发现 [1]。组蛋白是一类存在于真核细胞核内的碱性蛋白家族,是染色体的主要结构组分。它们富含赖氨酸和精氨酸残基,因此呈正电荷,能够紧密结合带负电的DNA,形成染色质。
2. 组蛋白的结构和类别
组蛋白主要分为两类:核心组蛋白和连接组蛋白。核心组蛋白包括四个成员:H2A、H2B、H3和H4。连接组蛋白包括H1或其同源物H5。在真核生物中,核心组蛋白通常在DNA复制过程中合成,达到S期时表达达到峰值。
所有核心组蛋白在从酵母到人类等各个物种中都表现出很强的保守性,它们具有一个涉及到组蛋白八聚体形成的C-末端组蛋白折叠域(HFD),以及伸出核心粒子并且可以接受各种翻译后修饰(PTMs)的无结构N-末端尾部。
在核小体内,核心组蛋白由两个(H3-H4)2四聚体与两个H2A-H2B二聚体组成,形成一个八聚体,周围绕着147bp的DNA被缠绕两次 [2]。H1与核小体的相互作用,以及在核小体入口/出口部位的额外DNA片段,导致了珠串状染色体的形成,并有助于建立更高阶的染色质结构 [3-5]。六个核小体,连同H1组蛋白,结合形成一个螺旋结构,随后绕在一个支架周围,再进一步缠绕以组装染色体基质。
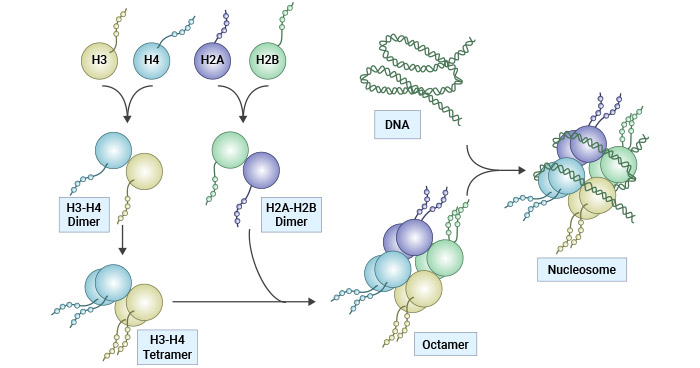
图1. 组蛋白结构和核小体组装
图片参考:https://www.nature.com/articles/cddis2014337
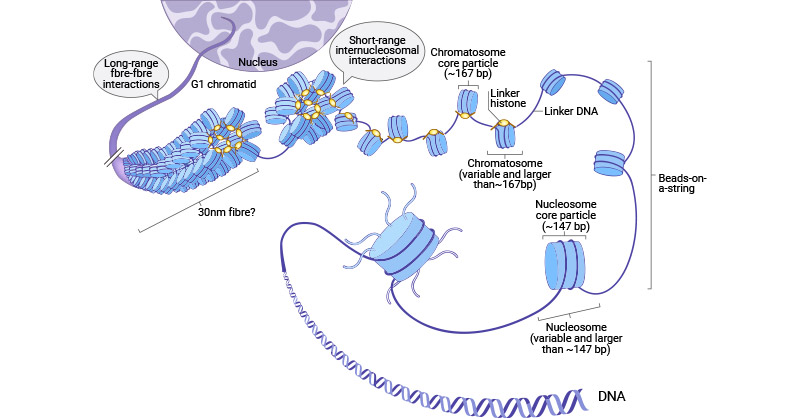
图2. 染色质折叠的多个层次和组蛋白结构
图片参考:https://www.nature.com/articles/cddis2014337
3. 组蛋白变异体
组蛋白变异体指的是组蛋白蛋白质,尤其是核心组蛋白的变异或稍微不同的形式。它们在结构上与核心组蛋白有相似之处,但具有独特的性质和功能。它们通过包裹更多或更少的DNA或改变核小体稳定性,为核小体赋予独特的结构和物理多样性。组蛋白变异体为真核细胞中染色质结构和表观遗传基因调控的复杂性和多样性做出贡献。
以下是人类组蛋白和变异体的列表:
| 超家族 | 家族 | 亚家族 | 成员 |
|---|---|---|---|
| Linker Histones | H1 | H1F | H1F0, H1FNT, H1FOO, H1FX |
| H1H1 | HIST1H1A, HIST1H1B, HIST1H1C, HIST1H1D, HIST1H1E, HIST1H1T | ||
| Core Histones | H2A | H2AF | H2AFB1, H2AFB2/3, H2AFJ, H2AFV, H2AFX, H2AFY, H2AFY2, H2AFZ |
| H2A1 | HIST1H2AA, HIST1H2AB/E, HIST1H2AC, HIST1H2AD, HIST1H2AG, HIST1H2AI, HIST1H2AJ, HIST1H2AK, HIST1H2AL, HIST1H2AM | ||
| H2A2 | HIST2H2AA3, HIST2H2AC | ||
| H2B | H2BF | H2BFM, H2BC12L, H2BFWT | |
| H2B1 | HIST1H2BA, HIST1H2BB, HIST1H2BC/E/F/G/I, HIST1H2BD, HIST1H2BH, HIST1H2BJ, HIST1H2BK, HIST1H2BL, HIST1H2BM, HIST1H2BN, HIST1H2BO | ||
| H2B2 | HIST2H2BE | ||
| H3 | H3A1 | HIST1H3A, HIST1H3B, HIST1H3C, HIST1H3D, HIST1H3E, HIST1H3F, HIST1H3G, HIST1H3H, HIST1H3I, HIST1H3J | |
| H3A2 | HIST2H3C | ||
| H3A3 | HIST3H3 | ||
| H4 | H41 | HIST1H4A, HIST1H4B, HIST1H4C, HIST1H4D, HIST1H4E, HIST1H4F, HIST1H4G, HIST1H4H, HIST1H4I, HIST1H4J, HIST1H4K, HIST1H4L | |
| H44 | HIST4H4 |
表格参考:维基百科
4. 组蛋白的功能
直到20世纪90年代初,组蛋白被认为仅是核DNA的包装材料,直到它们的调控功能被揭示。影响组蛋白功能的关键因素是位于组蛋白N-末端尾部附近的氨基酸侧链,这些侧链从核小体核心外部延伸,提供了对能够催化各种组蛋白后转录修饰(PTMs)的酶修饰机器的访问。组蛋白PTMs 控制了组蛋白蛋白的沉积和功能。组蛋白的功能主要体现在以下方面:
4.1 DNA包装和核小体形成
组蛋白负责将长链DNA包装和压缩成核小体,这有助于进一步紧缩DNA,并作为致密有序染色质的结构支架。这种紧缩使DNA适应细胞核,并防止DNA缠绕并保护免受损害。
4.2 基因调控和DNA复制
组蛋白的甲基化、乙酰化和磷酸化等修饰可以改变染色质结构,从而影响基因表达。例如,组蛋白甲基化可以抑制基因表达,而乙酰化与活跃转录有关。
4.3 表观遗传
组蛋白修饰的变化可以在细胞分裂期间传递给子细胞,促成表观遗传。表观遗传变化可以影响基因表达模式和细胞分化。
4.4 DNA修复
组蛋白参与DNA修复过程。特定的组蛋白变异体,如H2A.X,在DNA损伤部位迅速磷酸化,标记这些区域进行修复。
4.5 染色体分离
在细胞分裂期间,组蛋白在确保染色体准确分离到子细胞中起着作用。组蛋白的修饰可以影响染色体的凝聚和分离。
4.6 保持基因组完整性
组蛋白通过防止DNA暴露给有害物质和酶来保护基因组的完整性。
4.7 蛋白质结合的支架
组蛋白为与转录、DNA复制和修复过程有关的各种蛋白提供了一个支架。这些蛋白质可以与特定的组蛋白修饰相互作用,以执行它们的功能。
5. 组蛋白后翻译修饰(PTMs)
组蛋白主要在其N-末端尾部的氨基酸残基上进行后翻译修饰,通常在赖氨酸、精氨酸、丝氨酸、苏氨酸和酪氨酸残基上。这些修饰不会改变DNA序列,但可以调节染色质结构,从而调节基因表达。
常见的组蛋白修饰包括甲基化、乙酰化、磷酸化、SUMO化、泛素化和ADP-核糖基化。单一的组蛋白修饰通常不会独立发挥作用,而是在同一组蛋白上引导或抑制另一种修饰,形成一系列修饰的级联效应,这些修饰被特定一组蛋白识别,并随后被解释为特定染色质状态,以实现特定基因的调控。
这些不同组蛋白修饰之间的相互作用构成了一种标志或语言,也被称为“组蛋白密码”。组蛋白修饰可以影响所有与DNA相关的功能,包括基因转录、染色质组装、DNA修复和复制。这些修饰的影响取决于所添加的特定官能团以及发生修饰的确切位点。
参考文献:
[1] Kossel A, and Pringle H (1906). On protamines and histones [J]. H-S. Z. Physiol. Chem 49, 301–321.
[2] T.J. Richmond, C.A. Davey. The structure of DNA in the nucleosome core [J]. Nature, 423 (2003), pp. 145-150.
[3] P. Oudet, M. Gross-Bellard, P. Chambon. Electron microscopic and biochemical evidence that chromatin structure is a repeating unit [J]. Cell, 4 (1975), pp. 281-300.
[4] Thomas JO: Histone H1: location and role [J]. Curr Opin Cell Biol. 1999, 11: 312-317. 10.1016/S0955-0674(99)80042-8.
[5] Hergeth SP, Schneider R. The H1 linker histones: multifunctional proteins beyond the nucleosomal core particle [J]. EMBO Rep. 2015;16(11):1439–1453.





